
ORIGINAL ARTICLE
HDVR 2026, 37(1): 37-47
Exploring the relationship between DUSP3 expression and senescence-associated proliferative status
Emmanouil Chousakos1,3, Nikolaos Katsoulas1, Nikolaos I. Vlachogiannis2, Irene Theochari1, Nikolaos Kavantzas1, Alexandros Stratigos3, Andreas C. Lazaris1
11st Department of Pathology, Medical School, National and Kapodistrian University of Athens
21st Department of Propaedeutic Internal Medicine and Joint Rheumatology Program, Medical School, National and Kapodistrian University of Athens
31st Department of Dermatology-Venereology, “Andreas Syggros” Hospital, Medical School, National and Kapodistrian University of Athens
![]()
ABSTRACT
Background and Objectives: Dual specificity phosphatase 3 (DUSP3) regulates key components of the MAPK/ERK signaling pathway and has been implicated in melanocytic oncogenesis. Given the central role of oncogene-induced senescence in restraining melanocytic tumor progression, the relationship between DUSP3 expression and proliferative fitness remains unclear. This study aimed to investigate whether DUSP3 expression correlates with the proliferative status of melanocytic neoplasms.
Materials and Methods: A retrospective immunohistochemical analysis was performed on 172 melanocytic lesions, including common nevi, dysplastic nevi, and paired nevus and melanoma components of nevus-associated melanomas. β-galactosidase expression was used to stratify lesions according to proliferative fitness, and a biologically defined β-galactosidase–negative/low subgroup was analyzed separately.
Results: DUSP3 expression was signifi cantly higher in benign melanocytic lesions compared to melanomas, in both the entire cohort and the proliferative fitness subgroup. However, restriction of the analysis to lesions with retained proliferative capacity did not reveal significant differences in DUSP3 expression between them, either overall or within individual diagnostic categories. The relative expression gradient across lesion types remained consistent regardless of senescence status.
Conclusions: DUSP3 expression seems to not correlate with proliferative fitness or senescence status. These findings suggest that DUSP3 is not a central regulator of oncogene-induced senescence in melanocytic oncogenesis, and that its role is likely modulatory and context-dependent within broader signaling networks.
ΠΕΡΙΛΗΨΗ
Εισαγωγή και Στόχοι: Η φωσφατάση διπλής ειδικότητας 3 (DUSP3) ρυθμίζει κομβικά στοιχεία του σηματοδοτικού μονοπατιού MAPK/ERK, ενώ έχει συσχετιστεί με τη μελανοκυτταρική ογκογένεση. Δεδομένου δε του κρίσιμου ρόλου της ογκογονιδιακά επαγόμενης γήρανσης στον περιορισμό της εξέλιξης των μελανοκυτταρικών νεοπλασιών, η σχέση της έκφρασης της DUSP3 με τη διατηρημένη ικανότητα πολλαπλασιασμού παραμένει ασαφής. Σκοπός της μελέτης ήταν η διερεύνηση της συσχέτισης αυτής.
Υλικά και Μέθοδοι: Πραγματοποιήθηκε αναδρομική ανοσοϊστοχημική ανάλυση σε 172 μελανοκυτταρικές βλάβες, συμπεριλαμβανομένων κοινών σπίλων, δυσπλαστικών σπίλων και των τμημάτων του σπίλου και του μελανώματος από μελανώματα σχετιζόμενα με σπίλο. Η έκφραση της β-γαλακτοσιδάσης χρησιμοποιήθηκε για τη διαστρωμάτωση των βλαβών ως προς την πολλαπλασιαστική τους ικανότητα, ενώ αναλύθηκε ξεχωριστά υποομάδα με χαμηλή ή αρνητική έκφραση β-γαλακτοσιδάσης.
Αποτελέσματα: Η έκφραση της DUSP3 ήταν σημαντικά υψηλότερη στις καλοήθεις μελανοκυτταρικές βλάβες σε σύγκριση με τα μελανώματα, τόσο στο σύνολο του δείγματος όσο και στην υποομάδα με διατηρούμενη πολλαπλασιαστική ικανότητα. Ωστόσο, δεν αναδείχθηκαν στατιστικά σημαντικές διαφορές στην έκφραση της DUSP3 μεταξύ των δύο ομάδων, ούτε συνολικά ούτε εντός των επιμέρους διαγνωστικών κατηγοριών. Η διαβάθμιση της έκφρασης μεταξύ των διαγνωστικών κατηγοριών παρέμεινε σταθερή και στις δύο ομάδες.
Συμπεράσματα: Η έκφραση της DUSP3 φαίνεται πως δε συσχετίζεται με την πολλαπλασιαστική ικανότητα ή την κυτταρική γήρανση. Τα ευρήματα υποδηλώνουν ότι η DUSP3 δεν αποτελεί κεντρικό ρυθμιστή της ογκογονιδιακά επαγόμενης γήρανσης στη μελανοκυτταρική ογκογένεση, αλλά πιθανώς δρα με τροποποιητικό και εξαρτώμενο από το βιολογικό πλαίσιο τρόπο.
Keywords: Melanoma, nevi, skin neoplasms, melanocytic, oncogenesis, DUSP3, senescence.
Corresponding author: Emmanouil Chousakos, 1st Department of Pathology, Medical School, National and Kapodistrian University of Athens, 1st Department of Dermatology-Venereology, “Andreas Syggros” Hospital, Medical School, National and Kapodistrian University of Athens. Address: 40, Ypsilantou street, Athens 11521, Greece. Email: emmanouilchou@gmail.com
INTRODUCTION
Reversible phosphorylation, controlled through the coordinated actions of kinases and phosphatases, regulates numerous cellular processes and has become a central focus in cancer research. Disruptions in the balance of expression and/or activity between these opposing enzyme groups have been linked to the initiation and progression of many cancers, leading to abnormal phosphorylation of substrates that govern essential cellular functions, including signal transduction, growth, proliferation, differentiation, survival, DNA repair, and stress responses1,2. Several of these substrates are known targets of Dual Specificity Phosphatase 3 (DUSP3), underscoring its broad regulatory role across key oncogenic signaling pathways3-6.
DUSP3, also referred to as Vaccinia H1-related phosphatase, belongs to the atypical subgroup of the Dual Specificity Phosphatase family and acts on substrates within mitogen- and stress-activated signaling pathways, including ERK, JNK, and p38, thereby regulating cellular responses to external stimuli7. DUSP3-mediated δephosphorylation generally suppresses pathway activation and downstream signaling8,9. Beyond these functions, DUSP3 has also been implicated in controlling immune and inflammatory responses, as well as maintaining genomic repair and stability8,9. However, its limited substrate specificity, the diversity of its targets, and the often conflicting, context-dependent outcomes of its activity make it difficult to categorize DUSP3 strictly as either tumor-promoting or tumor-suppressive10. Although proper DUSP3 function appears to support cellular homeostasis and is frequently altered in various cancers, its role may shift depending on the underlying oncogenic drivers—such as mechanisms that bypass oncogene-induced senescence (OIS)—and the stage of tumor progression (benign, malignant, or metastatic)10-12. Consequently, both tumor-suppressive and tumor-promoting effects of DUSP3 have been observed across multiple cancer types, including cutaneous melanoma8-10.
Based on current evidence concerning the intracellular interactions occurring during melanocytic oncogenesis and DUSP3’s established substrates critically participating in it, multiple speculations have been raised supporting either its pro- or anti-tumoral profile10,13. DUSP3’s role in the oncogenesis of benign and malignant melanocytic tumors has been recently highlighted14, however its precise role in melanocytic oncogenesis and its relationship with cellular senescence remain unclear. OIS is recognized as an early tumor-suppressive barrier in melanocytic neoplasia, particularly in lesions driven by activating mutations in BRAF, most commonly BRAF V600E15. Constitutive BRAF signaling promotes an initial burst of proliferation in melanocytes but subsequently triggers a stable growth arrest characterized by cell-cycle inhibition, chromatin remodeling, and expression of senescence markers such as senescence-associated β-galactosidase16. This senescent state is thought to underlie the benign nature of most melanocytic nevi, where cells harbor oncogenic BRAF yet remain growth-restricted. Progression to malignant melanoma occurs when this senescence program is bypassed or disabled through additional genetic or epigenetic alterations or activation of alternative survival pathways, thereby restoring proliferative capacity and enabling tumor advancement17-19.
Given that melanocytic neoplasms are strongly influenced by OIS, and that several established DUSP3 substrates participate in senescence-associated signaling pathways during melanocytic oncogenesis, this study aimed to determine whether DUSP3 expression correlates with the proliferative status of neoplastic melanocytes, further elucidating the role of DUSP3 in melanocytic oncogenesis. Specifically, we analyzed variations in DUSP3 post-translational expression across different types of melanocytic neoplasms exhibiting varying degrees of atypia and distinct biological profiles, yet retaining proliferative capacity and lacking senescence features. The objective of this analysis was to explore the potential involvement of DUSP3 in the regulation or bypass of senescence mechanism.
MATERIALS AND METHODS
Data collection
This retrospective study received approval from the Bioethics Committee of the University of Athens. Cases of excised melanocytic neoplasms were identified through the biopsy archives of the 1st Department of Pathology, National and Kapodistrian University of Athens, and the Department of Pathology at Andreas Syggros Hospital. Inclusion criteria comprised: (i) histopathological diagnoses of common acquired melanocytic nevus (CN), dysplastic acquired melanocytic nevus (DN), irrespective of the degree of atypia, or cutaneous invasive nevus-associated melanoma (NAM); (ii) confirmation of these diagnoses and assessment of tissue suitability through independent review of hematoxylin and eosin–stained slides by two pathologists, including clear demarcation between nevus and melanoma areas in NAM specimens; (iii) availability of representative paraffin-embedded tissue blocks for further analysis; and (iv) sufficient accompanying demographic information. Lesions exhibiting spitzoid or congenital characteristics were excluded. Based on these criteria, four tissue subgroups were established: CN, DN, and the nevus and melanoma components of NAM (designated N-NAM and M-NAM, respectively). Although the latter two groups originated from the same NAM specimens, they were analyzed independently for study purposes. Patient sex, age at diagnosis, and histological subtype of nevi were also recorded.
Processing
Immunohistochemical staining for the markers of interest was carried out on 4-μm-thick formalin-fixed, paraffin-embedded tissue sections. Following overnight incubation at 37°C, deparaffinization, rehydration, and antigen retrieval were performed simultaneously using an automated PT Link system (Dako), with slides treated for 20 minutes at 96°C in EnVision FLEX Target Retrieval Solution High pH (50×) (Dako, Denmark). Endogenous peroxidase activity was blocked by incubation in 0.3% hydrogen peroxide prepared in Tris-buffered saline (TBS) for 15 minutes. After rinsing with TBS, sections were incubated for 1 hour with a rabbit polyclonal anti-DUSP3 antibody (LifeSpan Biosciences, Seattle, WA) at a 1:500 dilution. Parallel sections from the same lesions were also incubated for 1 hour with a mouse monoclonal anti-β-galactosidase antibody (clone OTI1E10; LifeSpan Biosciences) at a 1:200 dilution. Antibody detection was performed using a two-step polymer-based visualization system (goat anti-mouse/rabbit IgG–HRP; ImmunoLogic, WellMed BV, The Netherlands). Slides were first treated with a post-blocking solution for 30 minutes at room temperature, followed by incubation with the HRP-conjugated polymer for 30 minutes. Signal development was achieved using the EnVision FLEX HRP Magenta Substrate Chromogen System (Dako, Denmark). Finally, sections were counterstained with hematoxylin and mounted for evaluation.
Evaluation of immunohistochemical staining for DUSP3
Two independent observers (EC and NK) evaluated the immunohistochemical staining using light microscopy, assessing and documenting two distinct parameters for each case: (i) staining intensity within the examined lesion, graded as 0 (negative), 1 (weak), 2 (moderate), or 3 (strong); and (ii) staining extent, defined as the proportion of positively stained lesional cells, expressed as a percentage ranging from 0% to 100%. Any discrepancies between the reviewers were resolved through joint re-evaluation and consensus. Immunoreactivity was quantified using a modified Immunoreactive Score (IRS)20,21, previously applied in the assessment of DUSP2 expression in ovarian carcinoma22. For each specimen, a composite score was calculated by multiplying the intensity grade by the extent score. For this purpose, percentage values for staining extent were categorized as follows: 0% = 0, 1–25% = 1, 26–50% = 2, 51–75% = 3, and 76–100% = 4. Accordingly, each case was assigned to a positivity category based on the final calculated score as follows: scores of 0–1 were considered negative, 2–3 indicated mild positivity, 4–8 corresponded to moderate positivity, and 9–12 were classified as strong positivity20.
Evaluation of immunohistochemical staining for B-galactosidase
Two independent observers (EC and NK) evaluated the immunohistochemical staining using light microscopy, assessing and documenting staining intensity within the examined lesion, graded as 0 (negative), 1 (weak), 2 (moderate), or 3 (strong) for each case. Any discrepancies between the reviewers were resolved through joint re-evaluation and consensus.
Β-Galactosidase-Based stratification
Accordingly, from the total cohort, we isolated a biologically defined subgroup of lesions exhibiting low β-gal intensity (grade 0–1), hereafter referred to as the proliferative-fitness subgroup. This approach allowed focused evaluation of DUSP3 expression independently of advanced senescence-related metabolic alterations. Senescence-associated β-galactosidase (SA-β-gal) activity constitutes a well-established histochemical surrogate of oncogene-induced and replicative senescence in melanocytic neoplasms. Increased β-gal staining intensity reflects lysosomal expansion and metabolic remodeling characteristic of senescent, replication-incompetent melanocytes, whereas absent or weak staining is indicative of preserved proliferative fitness23. Given the pivotal role of senescence as a pivotal tumor-suppressive barrier in melanocytic oncogenesis, lesions exhibiting negative (grade 0) or weak (grade 1) β-gal staining were considered representative of melanocytic populations retaining proliferative competence. Representative immunohistochemical images are provided in Figure 1.
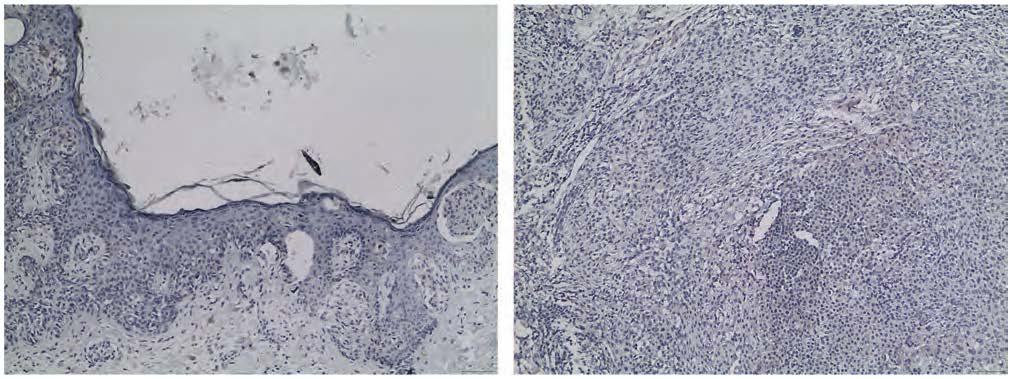
FIGURE 1. Images of the nevus component (N-NAM, left) and melanoma component (M-NAM, right) of the same NAM, weakly stained for β-gal. Both neoplasms were included in the proliferative fitness subgroup. 100x magnification.
Statistical analysis
Normality of the data was evaluated using the Shapiro–Wilk test. Continuous variables are reported as mean ± standard deviation (SD) for normally distributed data or as median with interquartile range (IQR) for non-normally distributed data, while categorical variables are expressed as absolute frequencies and percentages. For normally distributed variables, comparisons between two groups were performed using the independent samples t-test, with or without Welch’s correction as appropriate, whereas comparisons among three or more groups were conducted using one-way ANOVA followed by Dunnett’s post hoc test. For non-normally distributed variables, the Mann–Whitney U test was used for two-group comparisons and the Kruskal–Wallis test followed by Dunn’s multiple comparisons test for analyses involving three or more groups. All statistical tests were two-tailed, and analyses were performed using IBM SPSS Statistics version 28.0.
RESULTS
We conducted a retrospective analysis of 172 melanocytic neoplasm cases obtained from 130 patients, including 42 CN, 46 DN, and 84 paired samples representing the nevus and melanoma components of 42 NAMs. Of the 130 patients, 56 were male and 72 were female, with sex data unavailable for two individuals. The median age was 44 years (range 15–86; IQR 31 years). Among the 130 nevi evaluated, 88 (67.7%) were junctional, 39 (30.0%) were compound, and 3 (2.3%) were dermal. Detailed sample characteristics are summarized in Table 1.
Focusing on the proliferative fitness subgroup, 43 melanocytic lesions with retained proliferative competency from 39 patients were isolated from the entire sample, comprising 13 CN, 7 DN, 6 N-NAMs, and 17 M-NAMs. Among the 39 patients, 18 were male and 20 were female, with sex information unavailable for one individual. The overall median age at diagnosis was 55 years (range 18–85; IQR 37.5 years). When stratified by diagnosis, the median age was 36 years for CN (range 21–45; IQR 17.5), 65 years for DN (range 38–76; IQR 38), 62 years for N-NAMs (range 18–85; IQR 20), and 72 years for M-NAMs (range 18–85; IQR 25). Of the 26 nevi assessed for histologic subtype, 23 (88.5%) were junctional and 3 (11.5%) were mixed, while no dermal nevi were identified. Detailed characteristics are presented in Table 2.
Descriptive analysis of DUSP3’S expression for the entire sample
The immunohistochemical expression of DUSP3 was assessed using the IRS-based positivity classification (categorical), as previously described, which categorized staining into four levels: negative, mild, moderate, and strong. The findings are summarized in Table 3. Overall, nevi exhibited higher positivity than M-NAMs, with most CN, DN, and N-NAM cases showing moderate staining intensity (61.9%, 54.3%, and 57.1%, respectively). In contrast, the majority of M-NAMs displayed only mild positivity (52.4%). This pattern was also reflected in the mean IRS scores for each group, where benign melanocytic lesions demonstrated higher DUSP3 expression than malignant ones [mean (SD) IRS scores: CN 7.5 (3.5); DN 6.2 (3.6); N-NAMs 4.0 (2.9); M-NAMs 1.9 (1.1), p < 0.001]. Representative immunohistochemical images are provided in Figure 2. Negative staining was most frequently observed in M-NAMs and least frequently in CN, whereas strong staining was most common in CN and rarest in M-NAMs, with no M-NAMs showing strong positivity. Moreover, most NAMs (57.1%) arose from nevi with moderate positivity, while a notable proportion (21.4%) developed from negative lesions and only a small fraction (4.8%) originated from strongly positive nevi.
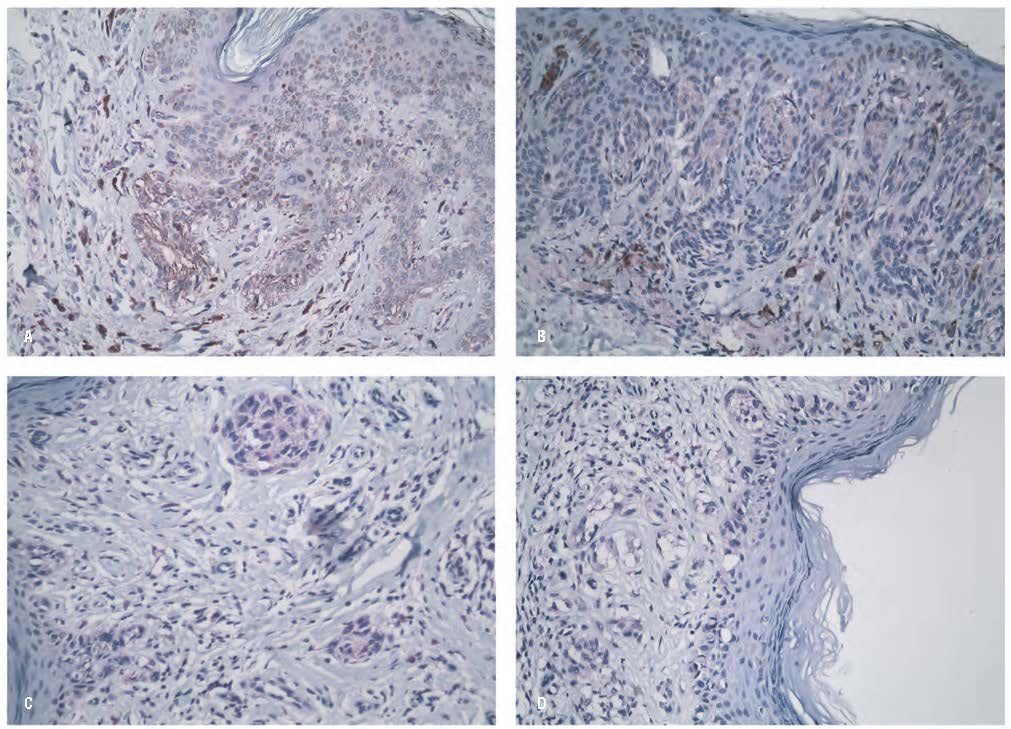
FIGURE 2. Images of lesions from the entire sample with IRS scores representative of their respective group means, immunohistochemically stained for DUSP3. 200x magnification. A: CN with DUSP3 IRS=8; B: DN with DUSP3 IRS=6; N-NAM with DUSP3 IRS=4; M-NAM with DUSP3 IRS=2.
Descriptive analysis of dusp3’s expression for the proliferative fitness subgroup
Within the β-gal–negative/low subgroup, DUSP3 expression was evaluated using the same IRS methodology applied to the entire cohort. The findings are summarized in Table 4. Overall, nevi (CN, DN, and N-NAMs) exhibited higher DUSP3 expression than M-NAMs. Specifically, most CN and DN cases showed moderate staining intensity (CN 46.2%, DN 28.6%), with a notable proportion of CN also showing strong positivity (30.8%). N-NAMs also largely demonstrated moderate positivity (66.7%). On the contrary, M-NAMs predominantly displayed negative staining (53.0%), as shown in Figure 3, and were frequently mildly positive (35.3%). This trend was mirrored in the mean IRS scores, where benign lesions had higher expression than malignant ones [mean (SD) IRS scores: CN 6.9 (4.2); DN 6.6 (4.6); N-NAMs 3.2 (1.3); M-NAMs 1.8 (1.3), p < 0.001]. Negative staining was most prevalent in M-NAMs and least common in CN, whereas strong staining was observed mainly in CN and was absent in M-NAMs.
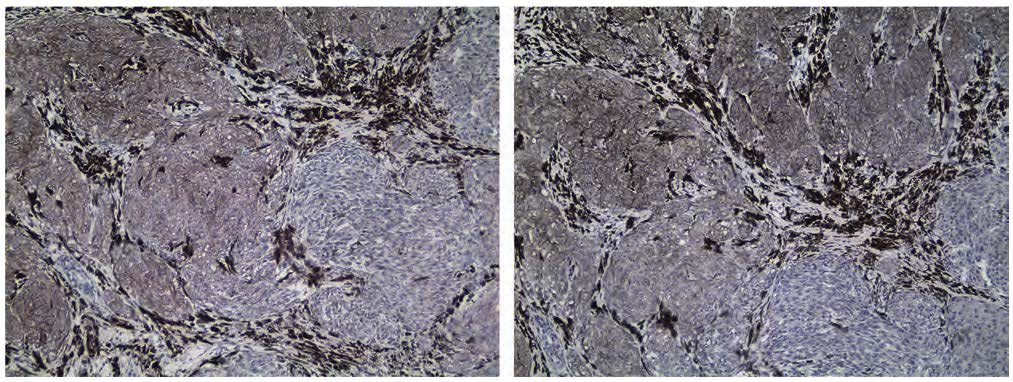
FIGURE 3. Characteristic image of an M-NAM from the proliferative fitness subgroup, negative for both β-gal (left) and DUSP3 (IRS=1) (right). 100x magnification.
Association of dusp3’s expression with proliferative fitness
Comparison of DUSP3 expression between the entire sample and the subgroup of tumors with retained proliferative fitness was performed. For each diagnostic category, mean numerical DUSP3 IRS scores of the proliferative fitness subgroup were compared with those of the corresponding full-sample group, as shown in Table 5. No statistically significant differences were observed for any diagnostic category (all p > 0.05). Considering the proliferative fitness subgroup as a whole, the mean IRS score was 4.33 ± 3.80, compared with 5.0 ± 3.6 in the entire cohort, a difference that also did not reach statistical significance (p > 0.05). These findings indicate that overall DUSP3 expression is not substantially altered when analysis is restricted to melanocytic lesions retaining proliferative fitness. Importantly, mean IRS values within each subgroup were directionally consistent with those of the entire cohort and showed no systematic upward or downward shifts.
DISCUSSION
OIS, particularly BRAF-induced OIS, represents a critical biological checkpoint that separates benign melanocytic proliferations from malignant transformation15. Constitutive activation of the MAPK/ERK pathway, most commonly driven by BRAF mutations, characterizes both benign and malignant melanocytic neoplasms and necessitates tight regulatory control to maintain cellular homeostasis. DUSP3 is thought to be among the negative regulators of this pathway, as it dephosphorylates and inhibits two key substrates within the cascade: the upstream receptor tyrosine kinases ERBB1/2 (EGFR1/2) and the terminal cytosolic effectors ERK1/23. Through this activity, DUSP3 is positioned to modulate both pathway initiation and downstream signaling progression, implicating it as a potential mediator of proliferation-senescence balance during melanocytic oncogenesis10. The observation that DUSP3 presence -and possibly functional competence- is more frequently associated with senescent or less hyperproliferative lesions, such as common and dysplastic nevi, than malignant melanocytic tissues suggests a putative protective role against malignant transformation14. Such an effect could reflect a compensatory response to oncogenic MAPK/ERK overactivation, contributing to appropriate cell-cycle regulation and maintenance of an OIS phenotype. Conversely, reduced DUSP3 expression might attenuate this oncosuppressive influence, facilitating escape from growth control and predisposing neoplastic populations to hyperproliferation, a potential early step toward melanoma development10.
However, the biological role of DUSP3 appears complex and potentially context-dependent. Experimental evidence indicates that DUSP3 is required for normal cell-cycle progression and mitosis, with its loss promoting senescence11. Consistently, stronger immunohistochemical expression has been documented in dysplastic nevi (lesions characterized by higher proliferative activity) compared to common nevi, as well as within their less mature, proliferative epidermal compartments of nevi24. These findings suggest that, while DUSP3 may physiologically regulate proliferation through modulation of MAPK/ERK signaling, its activity in the setting of sustained pathway hyperactivation could paradoxically interfere with the establishment or maintenance of OIS. Collectively, these observations raise the possibility that DUSP3 may influence melanocytic tumor biology not only by restraining oncogenic signaling but also by modulating the threshold for OIS induction or bypass.
Given these considerations, the present study aimed to further clarify the role of DUSP3 in melanocytic oncogenesis from the perspective of OIS regulation and maintenance, acknowledging its established importance in physiological cell-cycle control and its recently described potential oncosuppressive function14. We specifically investigated whether DUSP3 expression correlates with proliferative fitness across benign and malignant melanocytic lesions of various types.
Restriction of the analysis to lesions retaining proliferative competence, defined by absent or weak β-galactosidase activity, did not reveal significant differences in DUSP3 expression compared to lesions with variable proliferative status, either overall or within individual diagnostic categories. This lack of association argues against a direct mechanistic role for DUSP3 in the induction or maintenance of the senescent phenotype or in senescence-associated metabolic remodeling. The absence of divergence in DUSP3 expression among proliferatively competent lesions further indicates that DUSP3 levels do not adapt to the presence or absence of senescence. In other words, replicative status does not seem to influence DUSP3 expression regardless of the melanocytic neoplasm type. Instead, DUSP3 expression appeared more closely related to tumor type–specific biological characteristics rather than replicative status. Supporting this interpretation, the relative gradient of expression across diagnostic groups within the proliferative fitness subgroup mirrored that of the entire cohort, with nevi exhibiting higher levels and melanomas the lowest, independent of senescence markers.
Moreover, the identification of benign lesions (nevi) capable of sustained proliferation despite the putative protective barrier imposed by elevated DUSP3 expression suggests that melanocytic tumors may rely on DUSP3-independent mechanisms to maintain cell-cycle progression. This observation raises the possibility that alternative regulatory pathways compensate for increased DUSP3 activity, enabling a functional bypass of DUSP3-mediated control, or that the effects of DUSP3 vary in the context of benign oncogenesis, potentially facilitating OIS bypass17,25. In contrast, malignant lesions (NAMs) that retain proliferative capacity consistently exhibit reduced DUSP3 expression, supporting the notion that DUSP3 indeed exerts a protective role in malignant scenarios14.
To our knowledge, this is the first study to examine DUSP3 expression in relation to proliferative status in melanocytic oncogenesis. However, the analysis was limited to immunohistochemical assessment of DUSP3, without evaluation of its pre-translational regulation, including genetic alterations, epigenetic modifications, or transcriptional control, nor its specific functional activity. Additional limitations should be acknowledged. Our conclusions regarding melanoma primarily apply to the superficial spreading subtype, which is more commonly associated with preexisting nevi26,27. Although β-galactosidase is a robust and widely accepted marker of melanocytic senescence, reliance on a single marker represents an inherent limitation. Incorporation of additional markers would allow for more precise stratification of proliferative and senescent phenotypes. Moreover, integration of molecular and functional assays could further clarify the role of DUSP3 within senescence-associated signaling networks. Finally, additional studies are warranted to validate these findings by examining DUSP3 expression in well-defined cohorts of senescent melanocytic lesions.
In conclusion, while DUSP3 clearly participates in MAPK/ERK pathway regulation and physiological cell-cycle progression, our findings do not support a central or deterministic role for DUSP3 in governing OIS in melanocytic neoplasia. Rather, its contribution may be modulatory and context-dependent, embedded within broader, redundant signaling networks that collectively determine whether melanocytic cells undergo senescence or continue proliferating during tumor evolution.
CONFLICT OF INTERESTS STATEMENT
Authors declare no conflict of interests for this article.
Οι συγγραφείς δηλώνουν ότι δεν υπάρχει σύγκρουση συμφερόντων για το παρόν άρθρο.
FUNDING STATEMENT
This study is part of the doctoral (PhD) research of Emmanouil Chousakos (corresponding author), which has received
financial support from the A. G. Leventis Foundation’s Educational Grants Scheme.
Αυτή η μελέτη αποτελεί μέρος της διδακτορικής (PhD) έρευνας του Εμμανουήλ Χουσάκου (υπεύθυνος συγγραφέας), η οποία έχει λάβει οικο-
νομική υποστήριξη από το Πρόγραμμα Εκπαιδευτικών Υποτροφιών του Ιδρύματος Α. Γ. Λεβέντη.
BIBΛΙΟΓΡΑΦΙΑ
- Singh V, Ram M, Kumar R, Prasad R, Roy BK, Singh KK. Phosphorylation: Implications in Cancer. The Protein Journal. 2017 Jan 20;36(1):1–6.
- Stebbing J, Lit LC, Zhang H, Darrington RS, Melaiu O, Rudraraju B, et al. The regulatory roles of phosphatases in cancer. Oncogene [Internet]. 2014 Feb 1 [cited 2021 Jun 9];33(8):939–53. Available from: https://www.nature.com/articles/onc201380
- Todd JL, Tanner KG, Denu JM. Extracellular Regulated Kinases (ERK) 1 and ERK2 Are Authentic Substrates for the Dual-specificity Protein-tyrosine Phosphatase VHR. Journal of Biological Chemistry. 1999 May 7;274(19):13271–80.
- Amand M, Erpicum C, Bajou K, Cerignoli F, Blacher S, Martin M, et al. DUSP3/VHR is a pro-angiogenic atypical dual-specificity phosphatase. Molecular Cancer. 2014;13(1):108.
- Tambe MB, Narvi E, Kallio M. Reduced levels of Dusp3/Vhr phosphatase impair normal spindle bipolarity in an Erk1/2 activity-dependent manner. FEBS letters [Internet]. 2016 Aug;590(16):2757–67. Available from: https://pubmed.ncbi.nlm.nih.gov/27423135/
- Russo LC, Pault Y. M. Ferruzo, Forti FL. Nucleophosmin Protein Dephosphorylation by DUSP3 Is a Fine-Tuning Regulator of p53 Signaling to Maintain Genomic Stability. Frontiers in Cell and Developmental Biology. 2021 Mar 11;9.
- Pavic K, Duan G, Köhn M. VHR/DUSP3 phosphatase: structure, function and regulation. FEBS Journal. 2015 Apr 1;282(10):1871–90.
- Lucas Nascimento Monteiro, Minaya Y, Russo LC, Jessica Gadelha Farias, Fabio Luis Forti. DUSP3/VHR: A Druggable Dual Phosphatase for Human Diseases. Reviews of Physiology Biochemistry and Pharmacology. 2019 Jan 1;1–35.
- Russo LC, Farias JO, Ferruzo PYM, Monteiro LF, Forti FL. Revisiting the roles of VHR/DUSP3 phosphatase in human diseases. Clinics [Internet]. 2018 [cited 2026 Feb 4];73:e466s. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC6113852/
- Chousakos E, Katsoulas N, Kavantzas N, Stratigos A, Lazaris AC. The role of dual-specificity phosphatase 3 in melanocytic oncogenesis. Experimental dermatology [Internet]. 2022 Oct;31(10):1466–76. Available from: https://pubmed.ncbi.nlm.nih.gov/35899430/
- Rahmouni S, Cerignoli F, Alonso A, Tsutji T, Henkens R, Zhu C, et al. Loss of the VHR dual-specific phosphatase causes cell-cycle arrest and senescence. Nature cell biology [Internet]. 2006 May;8(5):524–31. Available from: https://pubmed.ncbi.nlm.nih.gov/16604064/
- Chen Y-R, Chou H-C, Yang C-H, Chen H-Y, Liu Y-W, Lin T-Y, et al. Deficiency in VHR/DUSP3, a suppressor of focal adhesion kinase, reveals its role in regulating cell adhesion and migration. Oncogene [Internet]. 2017 Autumn;36(47):6509–17. Available from: https://pubmed.ncbi.nlm.nih.gov/28759036/
- Joshi P, Seki T, Kitamura S, Bergano A, Lee B, Perera RJ. Transcriptome stability profiling using 5’-bromouridine IP chase (BRIC-seq) identifies novel and functional microRNA targets in human melanoma cells. RNA biology [Internet]. 2019 Oct;16(10):1355–63. Available from: https://pubmed.ncbi.nlm.nih.gov/31179855/
- Chousakos E, Katsoulas N, Vlachogiannis NI, Theochari I, Kavantzas N, Stratigos A, et al. The emerging role of dual specificity phosphatase 3 in melanocytic cancer: from oncogenesis to clinical value. Italian journal of dermatology and venereology [Internet]. 2025 Jun;160(3):195–203. Available from: https://pubmed.ncbi.nlm.nih.gov/40485572/
- Wajapeyee N, Serra RW, Zhu X, Mahalingam M, Green MR. Oncogenic BRAF Induces Senescence and Apoptosis through Pathways Mediated by the Secreted Protein IGFBP7. Cell. 2008 Feb;132(3):363–74.
- Michaloglou C, Vredeveld LCW, Soengas MS, Denoyelle C, Kuilman T, van der Horst CMAM, et al. BRAFE600-associated senescence-like cell cycle arrest of human naevi. Nature. 2005 Aug;436(7051):720–4.
- Constantinou SM, Bennett DC. Cell Senescence and the Genetics of Melanoma Development. Genes, chromosomes & cancer [Internet]. 2024 Oct;63(10):e23273. Available from: https://pubmed.ncbi.nlm.nih.gov/39422311/
- Lorbeer FK, Rieser G, Goel A, Wang M, Oh A, Yeh I, et al. Distinct senescence mechanisms restrain progression of dysplastic nevi. Furmanski P, editor. PNAS Nexus [Internet]. 2024 Feb 1 [cited 2025 Nov 20];3(2). Available from: https://academic.oup.com/pnasnexus/article/3/2/pgae041/7605632
- Ross AL, Sanchez MI, Grichnik JM. Nevus Senescence. ISRN Dermatology. 2011 Jun 22;2011:1–8.
- Fedchenko N, Reifenrath J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue – a review. Diagnostic Pathology. 2014 Nov 29;9(1).
- Remmele W, Stegner HE. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Der Pathologe [Internet]. 1987 May;8(3):138–40. Available from: https://pubmed.ncbi.nlm.nih.gov/3303008/
- Liu W, Tian X, Ding X, Zhang L. Expression of Dual-Specificity Phosphatase 2 (DUSP2) in Patients with Serous Ovarian Carcinoma and in SKOV3 and OVCAR3 Cells In Vitro. Medical science monitor: international medical journal of experimental and clinical research [Internet]. 2019;25:10180–9. Available from: https://pubmed.ncbi.nlm.nih.gov/31889045/
- Zimmermann T, Pommer M, Kluge V, Chiheb C, Muehlich S, Bosserhoff AK. Detection of Cellular Senescence in Human Primary Melanocytes and Malignant Melanoma Cells In Vitro. Cells [Internet]. 2022 Apr 28 [cited 2026 Feb 4];11(9):1489. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC9104908/
- Mitsui H, Kiecker F, Shemer A, Cannizzaro MV, Wang CQF, Gulati N, et al. Discrimination of Dysplastic Nevi from Common Melanocytic Nevi by Cellular and Molecular Criteria. Journal of Investigative Dermatology. 2016 Oct;136(10):2030–40.
- Wagner V, Gil J. An Epigenetic Switch: From Senescent Melanocytes to Malignant Melanoma (and Back). Cancer Cell. 2018 Feb;33(2):162–3.
- Pampena R, Lai M, Piana S, Lallas A, Pellacani G, Longo C. Nevus-associated melanoma: facts and controversies. Giornale italiano di dermatologia e venereologia: organo ufficiale, Societa italiana di dermatologia e sifilografia [Internet]. 2020 Feb;155(1):65–75. Available from: https://pubmed.ncbi.nlm.nih.gov/32100974/
- Bevona C. Cutaneous Melanomas Associated With Nevi. Archives of Dermatology. 2003 Dec 1;139(12):1620.